


80-81
Биомолекулы. Пептиды и белки
Свертывание белков
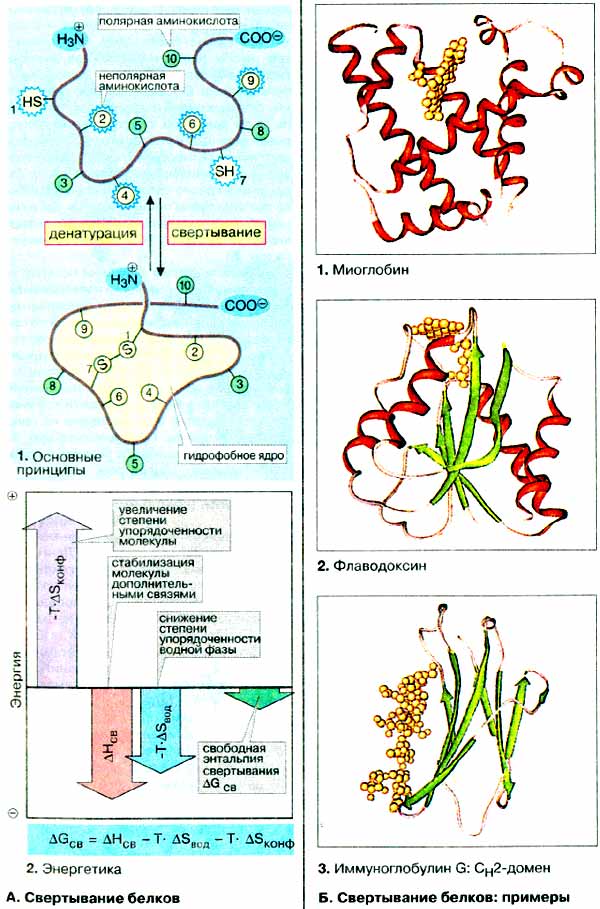 Информация относительно биологически
активной (нативной) конформации полипептидной цепи закодирована в аминокислотной
последовательности. Вторичные, третичные и четвертичные структуры многих белков
образуются в растворе самопроизвольно в пределах нескольких минут. Тем не менее
в клетке имеются специальные белки (шапероны, см. с. 230), функция которых
обеспечивать свертывание полипептидных цепей вновь синтезируемых белков.
Выяснение закономерностей свертывания полипептидных цепей является одной из
важнейших задач биохимии. В случае успеха появилась бы возможность предсказывать
нативные конформации белков на основании данных об аминокислотных
последовательностях, реконструируемых на основании относительно легко доступных
ДНК-последовательностей (см. c. 256).
Информация относительно биологически
активной (нативной) конформации полипептидной цепи закодирована в аминокислотной
последовательности. Вторичные, третичные и четвертичные структуры многих белков
образуются в растворе самопроизвольно в пределах нескольких минут. Тем не менее
в клетке имеются специальные белки (шапероны, см. с. 230), функция которых
обеспечивать свертывание полипептидных цепей вновь синтезируемых белков.
Выяснение закономерностей свертывания полипептидных цепей является одной из
важнейших задач биохимии. В случае успеха появилась бы возможность предсказывать
нативные конформации белков на основании данных об аминокислотных
последовательностях, реконструируемых на основании относительно легко доступных
ДНК-последовательностей (см. c. 256).
А. Свертывание белков
Свертывание полипептидной цепи в
нативную конформацию идет наиболее успешно в физиологических условиях. Потеря
нативной конформации, денатурация, наступает при экстремальных значениях
рН, высокой температуре или под действием органических растворителей,
детергентов и других денатурирующих веществ.
К факторам, стабилизирующим конформацию белка, относятся водородные
связи, дисульфидные мостики, электростатическое взаимодействие и комплексообразование
с ионами металлов (см. с. 73). Другим очень важным стабилизирующим
фактором является «гидрофобный эффект». Как отмечалось на с. 34,
в смеси неполярных вещесте с ведой происходит разделение фаз («эффект
масляных капель»), т. е. идет самопроизвольный процесс, при котором поверхность
контакта между фазами стремится быть минимальной. По аналогии с этим процессом
полипептидная цепь свертывается в водной среде таким образом, чтобы как можно
больше неполярных боковых групп аминокислотных остатков были бы спрятаны внутри
глобулы, тогда как полярные группы контактируют с водой (1).
Такой механизм позволяет объяснить распределение соответствующих группировок
и в молекуле инсулина (см. с. 83).
В настоящее время не существует полного описания энергетики
процесса свертывания полипептидной цепи (2).
В этом разделе обсуждается лишь предельно простая модель. В заданных условиях
конформация полипептидной цепи будет устойчивой лишь в том случае, если она
обладает минимумом свободной энергии (изменение свободной энергии свертывания
ΔGСВ имеет знак минус) (см. с. 22). Вместе
с тем свертывание полипептидной цепи повышает степень упорядоченности белковой
молекулы. А как указывалось на с. 26, рост упорядоченности
означает уменьшение энтропии системы (ΔSКОНФ — величина отрицательная),
а следовательно возрастание энтропийного члена в уравнении Гиббса-Гельмгольца
(-Т · ΔSКОНФ имеет знак плюс) (фиолетовая стрелка). На процесс
свертывания также оказывают стабилизирующее воздействие ковалентные и
другие типы связей, образующиеся в белковой глобуле. Поэтому изменение энтальпии
свертывания ΔHСВ — величина отрицательная (красная стрелка).
Другим фактором, влияющим на ход процесса, является изменение энтропии окружающей
среды (воды) за счет гидрофобного эффекта. При свертывании полипептидной цепи
снижается степень упорядоченности воды и образуется максимально возможное число
водородных связей. При этом возрастает энтропия водной среды, т. е. ΔSВОД
—величина положительная, а следовательно, энтропийный член уравнения -T · ΔSВОД
имеет знак минус (синяя стрелка). Таким образом, уменьшение энтропии полипептидной
цепи перекрывается ростом энтропии окружающей среды и энтропия системы в целом
возрастает. Следовательно, полипептидная цепь самопроизвольно принимает нативную
конформацию, характеризующуюся минимумом свободной энергии суммарной системы
(ΔGСВ — величина отрицательная) (зеленая стрелка).
Б. Свертывание белков: примеры
При сравнении наиболее крупных глобулярных белков становится
очевидным, что существует определенная схема свертывания полипептидной
цепи, которая воспроизводится с незначительными вариациями. Рассмотрим ряд примеров
(α-спирали выделены красным цветом, плоскости складчатого листа — зеленым),
глобулярные белки, построенные из α-спиралей, как например, миоглобин
(см. с. 330, гем выделен желтым цветом), встречаются
редко. Обычно наблюдаются сочетания складчатых листов и спирализованных участков,
как, например, во флаводоксине, небольшом флавопротеине (FMN выделен
желтым цветом), где 5 расположенных веером складчатых листов из пяти параллельных
тяжей формируют ядро молекулы; 4 α-спиральных участка окружают ядро снаружи.
Иммуноглобулин (см. с. 288) построен из нескольких
похожих доменов (независимых, компактно свернутых фрагментов полипептидной
цепи), в которых два антипараллельных складчатых листа из трех или четырех тяжей
образуют бочкообразную структуру (см. с. 292), Приведенный
на схеме СН2-домен несет олисахарид (желтый), который в более наглядной
форме приведен на с. 51.