


242-243
Молекулярная генетика
Созревание РНК
 Большинство меток организма содержит
полный набор генов, но обычно из этого набора используется крайне незначительный
объем информации. Постоянно транскрибируются только те гены, которые кодируют
структурные белки и ферменты промежуточного метаболизма. Кроме этих постоянно
необходимых генов имеется много других генов, активных только в определенных
типах клеток, при определенных метаболических условиях или во время
дифференцировки.
Большинство меток организма содержит
полный набор генов, но обычно из этого набора используется крайне незначительный
объем информации. Постоянно транскрибируются только те гены, которые кодируют
структурные белки и ферменты промежуточного метаболизма. Кроме этих постоянно
необходимых генов имеется много других генов, активных только в определенных
типах клеток, при определенных метаболических условиях или во время
дифференцировки.
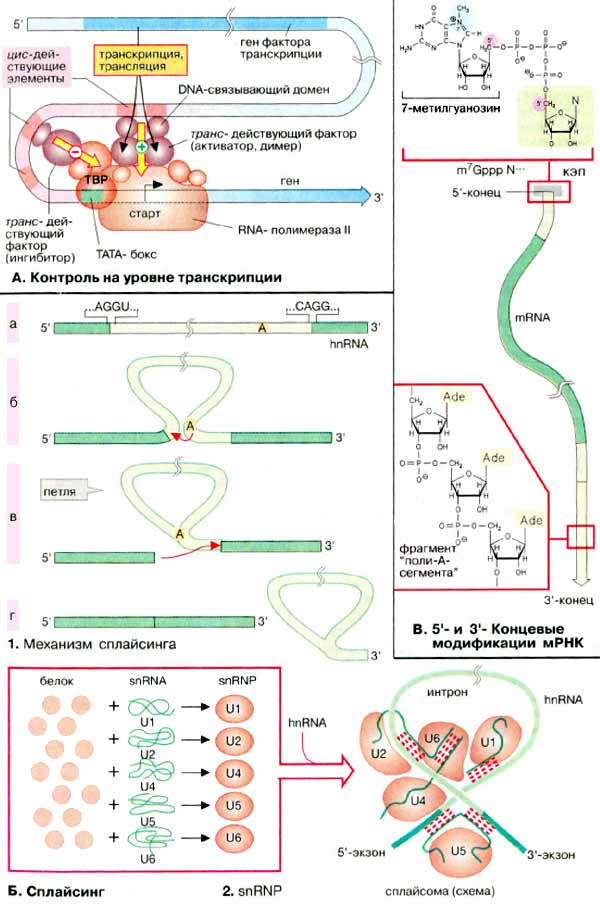
А. Контроль на уровне
транскрипции
Порядок транскрибирования генов
определяется регуляторной системой, которая носит название системы регуляции
транскрипции. Контроль транскрипции осуществляется структурами двух типов.
Большинство генов содержат в своем промоторном участке (см. с. 240)
несколько коротких сегментов ДНК (DNA) (регуляторные элементы,
цис-действующие элементы), с которыми могут связываться факторы
транскрипции. Регуляторные элементы, стимулирующие транскрипцию связанных с ними
генов, называются энхансерами (усилителями, от англ. enhancer).
Белки, подавляющие транскрипцию. — сайленсерами (успокоителями, от
англ. silencer). Факторы транскрипции — это белки, т. е. продукты других,
независимых генов. Поэтому их называют опосредованно действующими
факторами. Для процесса транскрипции генов требуются не только РНК-полимераза,
но и другие белки, называемые основными факторами транскрипции.
Установлено, что у эукариот таким фактором является ТАТА-связывающий
белок (ТСБ, англ. ТАТА-Вох Binding Protein, TBP), который
взаимодействует с основным регуляторным элементом. ТАТА-боксом, присутствующим в
большинстве генов (см. с. 240). С этим комплексом затем связываются другие
основные факторы транскрипции и РНК-полимеразы. Дополнительные факторы могут
влиять на инициацию транскрипции, связываясь с другими регуляторными
элементами. Отсюда они взаимодействуют с основным транскрипционным комплексом,
либо активируя, либо ингибируя его. Такие факторы активируют, например,
комплексы стероидных гормонов с рецепторами (см. с. 366). По завершении
транскрипции из гяРНК вырезаются интроны (см. с. 240), содержащие некодирующие
последовательности.
Б. Сплайсинг
Сплайсинг РНК катализируется комплексами белков с РНК, известными
как «малые ядерные рибонуклеопротеидные частицы» (мяРНП, англ. small
nuclear ribonucleic particles, snRNP). Интроны, входящие в гяРНК (hnRNA),
имеют специфические последовательности на 3'- и 5'-концах (а).
На первой стадии сплайсинга ОН-группа аденозилового остатка, расположенного
в интроне, атакует (при участии мяРНП) и расщепляет фосфодиэфирную связь на
5'-конце интрона (б). Одновременно в интроне образуется
новая связь, которая придает ему форму петли (в).
На второй стадии терминальная ОН-группа 5'-концевого интрона атакует связь в
3'-конце интрона. В результате оба экзона соединяются, а интрон освобождается
(г).
В этой реакции принимают участие пять
различных мяРНП (U1, U2, U4, U5 и U6). В каждой из реакций
задействованы несколько белковых молекул и одна молекула мяРНК
(snRNA) (см. с. 88). Во время сплайсинга комплексы из гяРНК и мяРНП образуют
сплайсому. Полагают, что мяРНК в сплайсоме образуют канонические пары
друг с другом и с гяРНК и таким образом фиксируют и ориентируют их реакционные
группы. Собственно катализ обусловлен РНК-составляющей сплайсомы Такие
каталитические РНК носят название рибозимов.
В. 5'- и 3'-Концевые модификации
мРНК
У эукариот после завершения собственно
транскрипции 5'-конец растущей молекулы РНК блокируется структурой, которая
называется кэп (от англ. cap). В случае мРНК кэп состоит из 7'-метил-ГТФ
и защищает РНК от гидролиза 5'-экзонуклеазами. В конце транскрипции к 3'-концу
присоединяется полиадениловая последовательность, которая может включать
до 200 звеньев АМФ (AMP). Только после этого созревшая мРНК (mRNA) покидает
ядро.