


238-239
Молекулярная генетика
Репликация
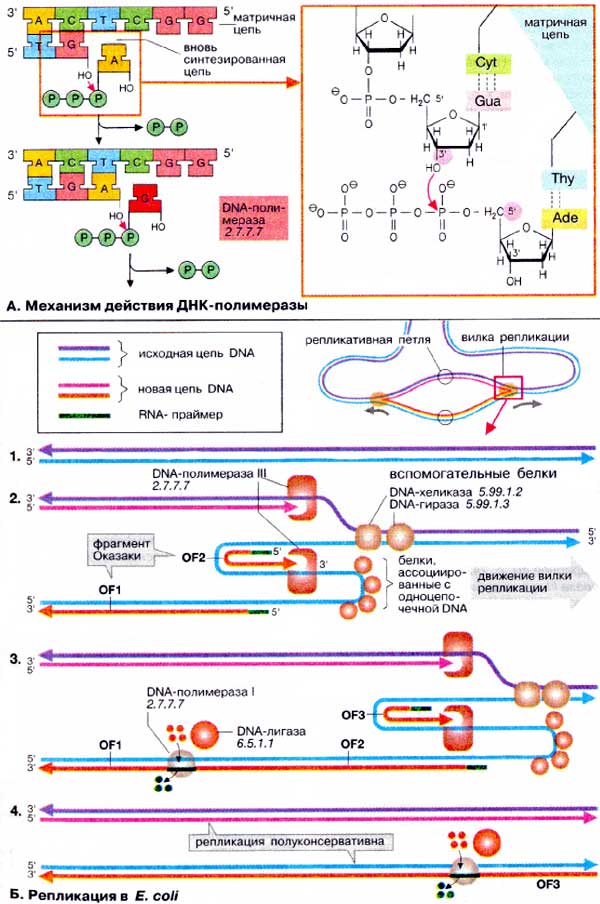 А. Механизм действия ДНК-полимеразы
А. Механизм действия ДНК-полимеразы
Для передачи дочерним клеткам
генетической информации в процессе репликации ДНК (DNA) должна быть создана
копия генома. Репликация ДНК осуществляется ДНК-зависимыми ДНК-полимеразами.
Эти ферменты используют в качестве шаблона одну из цепей двойной спирали
ДНК, так называемую матрицу. На матрице, начиная с короткой стартовой
последовательности (праймера), ферменты синтезируют комплементарную цепь
и воспроизводят в итоге исходную двухтяжевую ДНК. Субстратами ДНК-полимераз
являются четыре дезоксирибонуклеотидтрифосфата: аденозин-, гуанозин-,
тимидин- и цитозинтрифосфаты. При каждом шаге синтеза ДНК происходит
спаривание нуклеотида с соответствующим азотистым основанием матричной цепи.
Затем α-фосфатная группа связанного нуклеотида подвергается нуклеофильной атаке
со стороны 3'-ОН-группы предыдущего нуклеотида. За этим следует удаление
дифосфата и образование новой фосфодиэфирной связи. Эти этапы повторяются снова
и снова по мере движения ДНК-полимеразы от одного основания к следующему вдоль
матрицы. В соответствии с этим механизмом матричная цепь ДНК считывается в
направлении 3'→5'.
В большинстве клеток имеется несколько
ДНК-полимераз. Наряду с ферментами, которые осуществляют собственно репликацию,
существуют полимеразы, которые включены в процессы репарации ДНК (см. с. 252)
или реплицируют митохондриальную ДНК эукариот. Большинство ДНК-полимераз
построены из множества субъединиц, роль которых до конца не
выяснена.
Б. Репликация в Е.
coli
В настоящее время процесс репликации у
прокариот достаточно изучен, в то время как многие аспекты эукариотической
репликации остаются неясными. Однако с большой долей вероятности можно
утверждать, что в большинстве клеток этот процесс протекает в основном
одинаково. На схеме показана простейшая схема репликации у бактерии
Escherichia coli. В бактериях репликация начинается со специфической
точки в кольцевой ДНК (область начала репликации) и продолжается в обоих
направлениях. В результате образуются две репликативные вилки, которые
продвигаются в противоположных направлениях, т. е. обе цепи реплицируются
одновременно. На схеме исходная ДНК (1) окрашена в голубой и фиолетовый цвета, а
вновь синтезирующаяся — в розовый и оранжевый. В функционировании каждой вилки
принимают участие множество различных белков, из которых здесь указаны наиболее
важные.
Каждая репликативная вилка (2)
включает по крайней мере две молекулы ДНК-полимеразы III, ассоциированные
с несколькими вспомогательными белками. К последним относятся ДНК-топоизомеразы
(гиразы), которые раскручивают плотно свернутую двойную спираль ДНК, и хеликазы,
которые расплетают двухтяжевую ДНК на две цепи. Поскольку матричная цепь
всегда читается в направлении 3'→5' (см. выше), только одна из цепей может
считываться непрерывно (розовая/фиолетовая; 2).
Другая цепь (голубого цвета) считывается в направлении, противоположном движению
репликативной вилки. В результате на матрице вначале синтезируются короткие
фрагменты новой цепи ДНК (зеленый/оранжевый), так называемые фрагменты Оказаки
(OF), названные так по имени их первооткрывателя. Каждый фрагмент начинается
с короткой РНК-затравки (праймера, зеленого цвета), необходимой для функционирования
ДНК-полимеразы. Праймер синтезируется специальной РНК-полимеразой («праймаза»,
на схеме не показана), ДНК-полимераза III достраивает этот праймер до
фрагмента ДНК длиной 1000-2000 дезоксинуклеотидных звеньев (оранжевого цвета).
Синтез этого фрагмента далее прерывается, и новый синтез начинается со следующего
РНК-праймера. Индивидуальные фрагменты Оказаки первоначально не связаны друг
с другом и все еще имеют РНК на 5'-концах (3).
На некотором расстоянии от репликативной вилки ДНК-полимераза I начинает
замещать РНК-праймер последовательностью ДНК. В завершение остающиеся одноцепочечные
разрывы репарируются ДНК-лигазой. В образованной таким образом двойной
спирали ДНК только одна из цепей синтезирована заново. Поэтому говорят, что
репликация ДНК происходит по полуконсервативному механизму.